组蛋白甲基化通常发生在 H3 和 H4 的精氨酸 (Arg 或 R) 和赖氨酸 (Lys 或 K) 残基上。这些精氨酸和赖氨酸都可以被单甲基化或二甲基化,赖氨酸还能再被三甲基化。组蛋白的甲基化修饰受到组蛋白甲基转移酶 (HMTs) 和组蛋白脱甲基化酶 (HDMs)调控。但与乙酰化修饰的生物学效应不同,甲基化后组蛋白赖氨酸残基可以激活或抑制基因转录,这取决于具体的情况 (如甲基化的位点,状态等),例如 H3K4me2/3, H3K36me1/3, H3K79me1/2 和 H4K20me1 与转录激活相关,而 H3K9me2/3, H3K27me2/3, H3K79me3 和 H4K20me3 与转录抑制相关。
写入: 组蛋白甲基转移酶 (HMTs)
组蛋白甲基转移酶 (HMTs) 分为两类:赖氨酸甲基转移酶 (KMTs) 和精氨酸甲基转移酶 (PRMTs)。
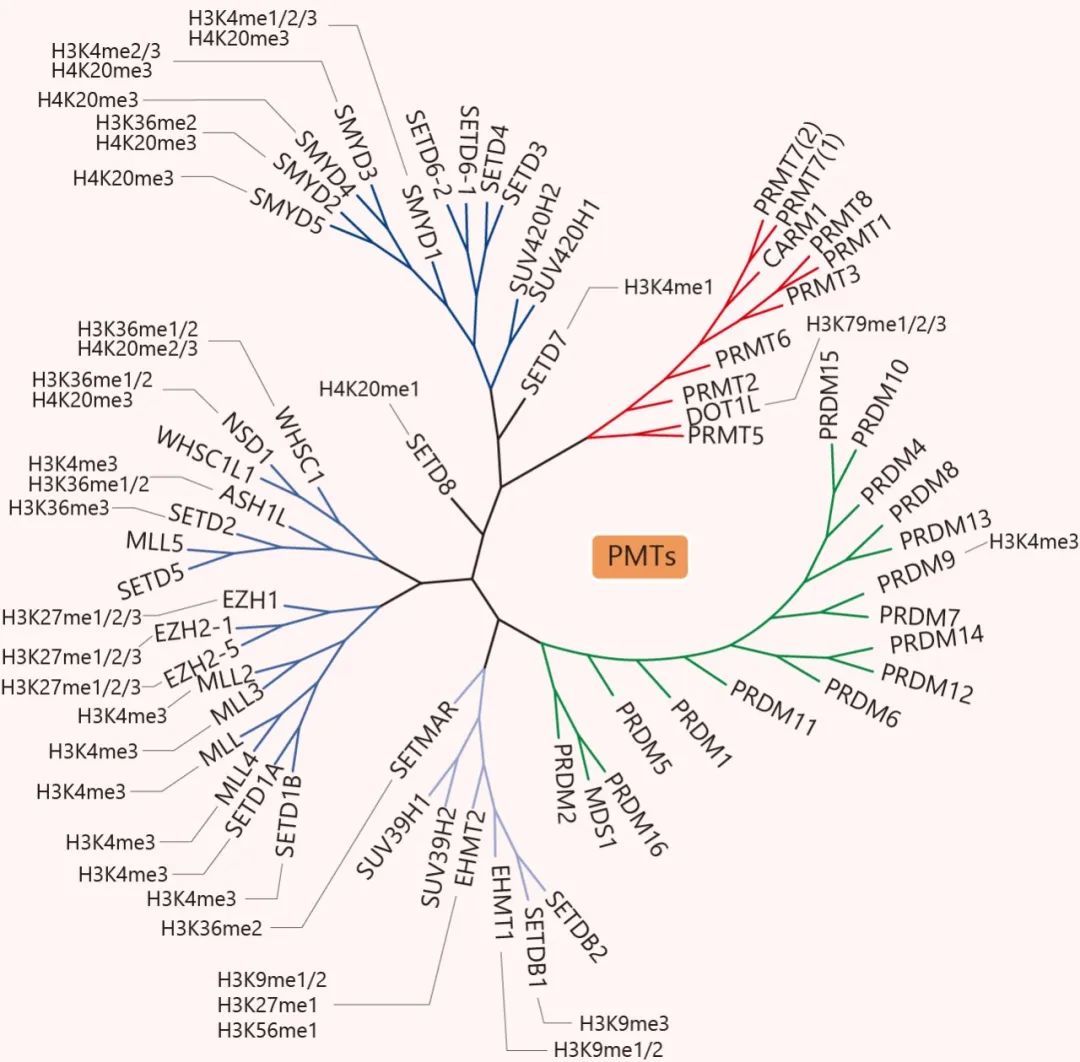
KMTs 根据催化结构域序列,可分为含 SET 结构域和非 SET 结构域。SET 结构域是组蛋白甲基转移酶的重要结构域,也是大多数转移酶含有的结构域,负责甲基转移酶的酶促活性,包括 SUV39, SET1, SET2, EZH (著名的 EZH2 就在这个家族啦, 可对 H3K27 进行单,二和三甲基化),RIZ (PRDM, SMYD, SUV420) 等家族。而不含 SET 结构域的蛋白较少,如 DOT1L 蛋白。DOT1L 是已知的靶向组蛋白 H3K79 位置的组蛋白甲基转移酶。H3K79 位于组蛋白 H3 的球状结构域中,但它暴露在核小体表面上,在这里它可以被 DOT1L 甲基化。因此,DOT1L 的催化发生在核小体表面而不是 N 末端尾巴上。
PRMTs 根据其催化活性可分为三类,催化精氨酸的单甲基化 (MMA),不对称 (ADMA) 或对称二甲基化 (SDMA)。I 型 PRMTs (PRMT1, PRMT2, PRMT3, PRMT4, PRMT6 和 PRMT8) 产生单或不对称二甲基化精氨酸 (ADMA),II 型 PRMTs (PRMT5 和 PRMT9) 产生单或对称二甲基化精氨酸 (SDMA)。而 Ⅲ 型的 PRMT7,只产生 MMA。组蛋白甲基化转移酶的成员实在不少,并有各自的识别位点,就列在树上给小伙伴们看吧。
擦除: 组蛋白脱甲基酶 (HDMs)
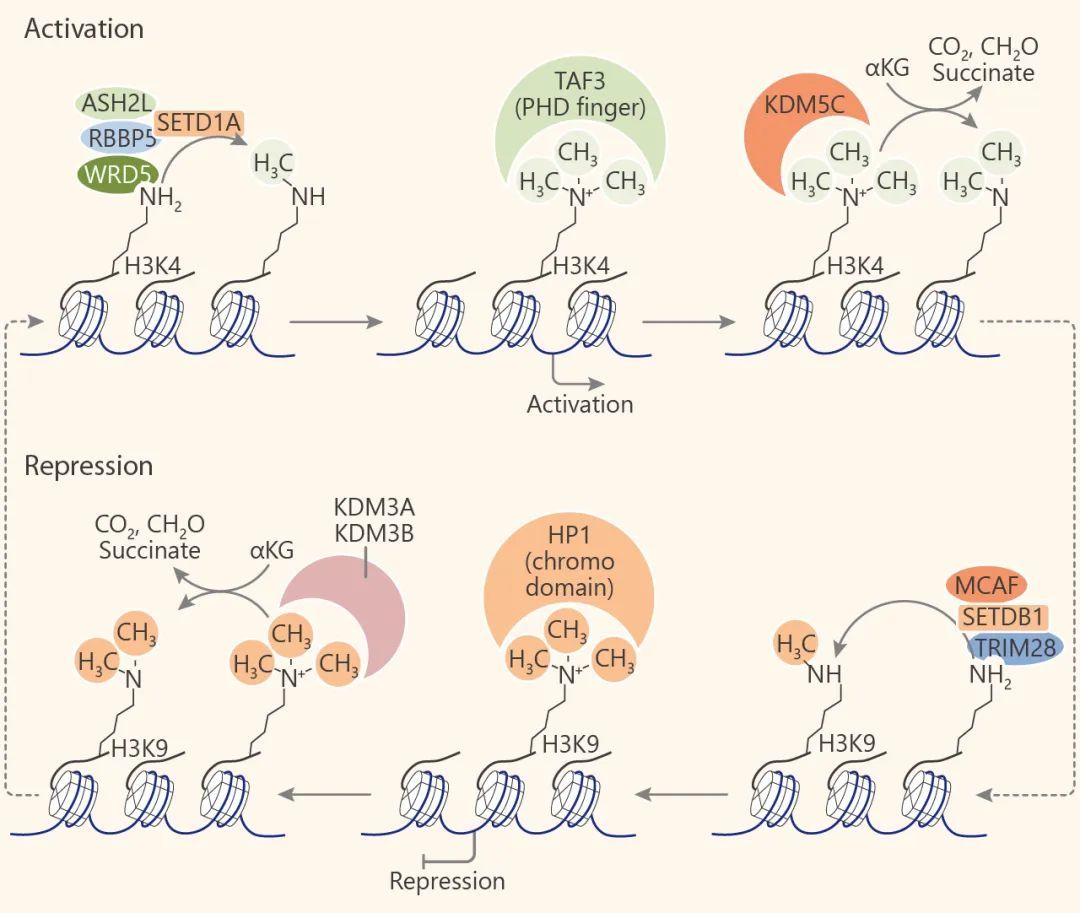
组蛋白去甲基化酶与转移酶的作用相反。组蛋白去甲基酶既能靶向组蛋白又能靶向非组蛋白底物。目前已经鉴定出两个进化上保守的组蛋白去甲基化酶家族:赖氨酸特异性去甲基化酶 (LSD)和Jumonji C (JMJC)蛋白家族,它们利用不同的反应机理来去甲基。LSD 蛋白家族由 LSD1 和 LSD2 组成,它们通过 FAD 依赖的胺氧化反应 (flavin adenine dinucleotide-dependent amineoxidase) 对单和二甲基化的赖氨酸残基进行脱甲基化。其中 LSD1 (KDM1A) 是第一个发现的组蛋白赖氨酸脱甲基酶 (KDM), 催化 H3K4me1/2, H3K9me1/2 的脱甲基化,另外,LSD1 也可以对非组蛋白脱甲基化,如 p53 上的 K370me1 和 K370me2, DNMT1 上的 Lys1096 和 E2F1 上的 Lys185。
而 JMJC 家族催化的脱甲基酶反应是一种依赖于铁 (II) 和 α-酮戊二酸的双加氧酶 (Fe(II)/α-ketoglutarate-dependent hydroxylase) 反应。这使得 JMJC 家族的酶与 LSD 家族不同,能够脱三甲基赖氨酸残基。JMJC 家族由 30 个成员组成,目前为止,这些成员中的 18 个已显示具有组蛋白脱甲基酶活性。因为组蛋白脱甲基化酶的成员也实在不少,就不一一列举了,小伙伴们可以参考这个表。
读取:组蛋白甲基化的识别蛋白
甲基化组蛋白的识别是通过具有甲基结合域的蛋白来实现的,这些结合域包括 ADD, Ankyrin, BAH, Chromobarrel, Chromodomain, Double Chromodomain (DCD), MBT, PHD, PWWP, TTD, Tudor, WD40 以及 zf-CW。其中含 Chromodomain, MBT 重复序列,PWWP, Tudor, DCD 蛋白组成了 Royal 超家族。这些域是怎么识别组蛋白甲基化位点呢?许多研究表明,含 Chromodomain 蛋白的 HP1 和 Chd1,分别可以识别 H3K9me 和 H3K27me。而人的 L3MBTL1 蛋白,是已知的转录阻遏物,以严格依赖组蛋白甲基化标记的方式 (如 H4K20me1/2 和 H1K26me1/2) 压紧染色质。L3MBTL1 含有三个 MBT 域,都很重要。例如它的第二个 MBT 结构域对 H1K26me1/2 和 H4K20me1/2 的结合很重要。含有 PHD 手指基序的 BPTF, RAG2, PYGO 和 ING2 都可以识别并结合到 H3K4me3。另外,含 WD40 重复序列的蛋白如 WDR5,可通过与 MLL, RBBP5, ASH2L 和 DPY30 形成蛋白质复合物,促进组蛋白 H3K4 甲基化。
如同武林高手组合出招一样,组蛋白甲基化的不同位点和模式可以演化出很多的甲基化修饰模式,增加了受组蛋白甲基化调节的基因表达的复杂性和多样性。而 HMTs 和 HDMs 小心地维持着组蛋白甲基化的水平,因而也就不难理解它们的失调与癌症之间密切的关系,如组蛋白甲基转移酶 NSD1 和 EZH2 在许多肿瘤中过表达,DOT1L 在白血病中有着广泛的作用等等。组蛋白脱甲基酶 KDM1A, KDM5B 分别在低分化神经母细胞瘤和前列腺癌中过表达。LSD1 与 p53 的直接相互作用会降低 p53 的活性,包括 p21 的表达降低,与肿瘤发生有关等等。另外,对组蛋白甲基化标记的误读 (组蛋白甲基化的读取蛋白活性异常) 也与许多人类疾病有关,包括发育异常以及癌症。因此,这些蛋白质的小分子抑制剂是有用的化学探针或潜在的治疗剂。
部分靶向组蛋白甲基化修饰的蛋白抑制剂
|
化合物 |
作用 |
|---|---|
|
HMTs |
|
|
Tazemetostat |
选择性,具有口服活性的 EZH2 抑制剂,用于治疗上皮样肉瘤;抑制含有 PRC2 复合体的野生型 EZH2 的活性; FDA Approved |
|
GSK126 |
有效的,选择性的 EZH2 甲基转移酶抑制剂;在体内外明显抑制肿瘤形成,降低细胞迁移,侵袭,逆转耐药 |
|
AZ505 |
有效的,具有选择性的 SMYD2 抑制剂 |
|
Pinometostat |
有效的 DOT1L 组蛋白甲基转移酶抑制剂; Phase 2 |
|
EPZ015666EPZ015666 |
有口服活性的 PRMT5 抑制剂 |
|
MS023 |
有效的,选择性的,具有细胞活性的人 I 型蛋白精氨酸甲基转移酶 (PRMTs) 抑制剂 |
|
HDMs |
|
|
GSK-J4 |
有效的 H3K27me3/me2 脱甲基化酶 JMJD3/KDM6B 和 UTX/KDM6A 双抑制剂 |
|
GSK2879552 |
具有口服活性的,不可逆的 LSD1 抑制剂,具有抗肿瘤活性; Phase 1 |
|
Seclidemstat |
有效的 LSD1 抑制剂; Phase 1 |
|
JIB-04 |
是 Jumonji 组蛋白脱甲基酶广谱抑制剂 |
|
Reader |
|
|
UNC 669 |
L3MBTL1 和 L3MBTL3 的抑制剂 |
MCE 的所有产品仅用作科学研究,我们不为任何个人用途提供产品和服务
HY-L005 表观遗传化合物库 Epigenetics Compound Library
缩写:
HMTs: histone methyltransferases
HDMs: histone demethylases
PHD: plant homeodomain
PRMTs: protein arginine methyltransferase
KMTs: histone lysine methyltransferases
DOT1L: Dot1-Like histone methyltransferase
MMA: monomethylarginine
ADMA: asymmetric dimethylarginine
SDMA: symmetric dimethylation
LSD: lysine-specific histone demethylase
KDM: histone lysine demethylases
HP1: heterochromatin protein 1
参考文献
1. Cheng Y, et al. Targeting epigenetic regulators for cancer therapy: mechanisms and advances in clinical trials. Signal Transduct Target Ther. 2019 Dec 17;4:62.
2. Jambhekar A, et al. Roles and regulation of histone methylation in animal development. Nat Rev Mol Cell Biol. 2019 Oct;20(10):625-641.
3. Xin Yi, et al. Histone methyltransferases: novel targets for tumor and developmental defects. Am J Transl Res. 2015 Nov 15;7(11):2159-75.
4. Tian X, et al. Histone lysine-specific methyltransferases and demethylases in carcinogenesis: new targets for cancer therapy and prevention. Curr Cancer Drug Targets. 2013 Jun;13(5):558-79.
5. Michalak EM, et al. s of DNA, RNA and histone methylation in ageing and cancer. Nat Rev Mol Cell Biol. 2019 Oct;20(10):573-589.
6. Kooistra SM, et. Molecular mechanisms and potential functions of histone demethylases. Nat Rev Mol Cell Biol. 2012 Apr 4;13(5):297-311.
7. Milite C, et al. The emerging role of lysine methyltransferase SETD8 in human diseases. Clin Epigenetics. 2016 Sep 22;8:102.
8. Islam AB, et al. Co-regulation of histone-modifying enzymes in cancer. PLoS One. 2011;6(8):e24023.
9. Huang J, et al. p53 is regulated by the lysine demethylase LSD1. Nature. 2007 Sep 6;449(7158):105-8.
10. Musselman CA, et al. Perceiving the epigenetic landscape through histone readers. Nat Struct Mol Biol. 2012 Dec;19(12):1218-27.
11. Zhu H, et al. Molecules. 2020 Jan 29;25(3). pii: E578.Small Molecules Targeting the Specific Domains of Histone-Mark Readers in Cancer Therapy.
12. Albert M, et al. Histone methyltransferases in cancer. Semin Cell Dev Biol. 2010 Apr;21(2):209-20.
13. D’Oto A, et al. Histone demethylases and their roles in cancer epigenetics. J Med Oncol Ther. 2016;1(2):34-40.
14. Arrowsmith CH, et al. Epigenetic protein families: a new frontier for drug discovery. Nat Rev Drug Discov. 2012 Apr 13;11(5):384-400.
15. McCabe MT, et al. Targeting Histone Methylation in Cancer. Cancer J. 2017 Sep/Oct;23(5):292-301.
16. Magliulo D, et al. Lysine-Specific Demethylase 1A as a Promising Target in Acute Myeloid Leukemia.Front Oncol. 2018 Jul 19;8:255